Parte 9:Fósseis Transicionais
"Quanto à origem do ser humano, este é o aspecto mais controverso da Teoria Geral da Evolução. O que revelam os fósseis? Henry Gee, escritor-chefe de ciência da revista Nature, é muito pessimista a respeito do assunto: 'Nenhum fóssil é enterrado com o seu registro de nascimento... os intervalos de tempo que separam os fósseis são tão imensos que nós não podemos dizer nada definitivo sobre sua possível conexão através da ancestralidade ou descendência...[cada fóssil] é um ponto isolado, com nenhuma conexão conhecida com qualquer outro fóssil dado, e tudo flutua num irresistível mar de lacunas... entre 10 e 5 milhões de anos atrás - diversos milhares de gerações de criaturas - tudo isso cabe dentro de uma pequena caixa...Pegar uma linhagem de fósseis e afirmar que eles representam uma linhagem não é uma hipótese científica que possa ser testada, mas uma afirmação que tem a mesma validade como uma estória para dormir - interessante, talvez até instrutiva, mas não é científica'.['In Search of Deep Time: Beyond the Fossil Record to a NewHistory of Life', New York: The Free Press, 1999]."
------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------------
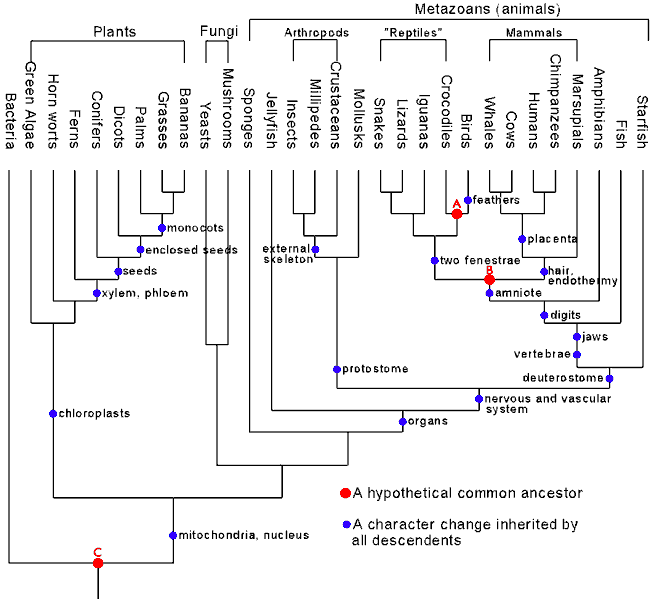
Todos os argumentos que tentam fazer acreditar que um algum evolucionista não crê que o registro fóssil pode evidenciar a genealogia da vida, baseia-se em desconhecimento sobre a metodologia da sistemática filogenética.No caso acima, há de se esclarecer que Gee não diz ou sequer deixa subentendido que está pessimista a utilização de fósseis como evidência de ancestralidade comum.Isso porque espécies fósseis são utilizados e ajudam a sustentar as árvores filogenéticas, mas são interpretados como ponta de ramos extintos, não como ancestrais.O fato que fósseis transicionais não é sinônimo de fósseis ancestrais é conhecido desde o século XIX, porque para a maioria dos evolucionistas "forma ancestral" é apenas uma inferência aproximada. Mesmo uma transição de espécie nem sempre é um ancestral, já que frequentemente não há como distinguir um ancestral de uma ponta de ramo extinta estreitamente aparentada ao ancestral (Darwin,1987.p.249).Ainda assim, uma forma transicional pode representar a morfologia predita de um ancestral comum ou de um ancestral recente,coisa vista nos fósseis de Australopithecus e de Homo.Porque mais de uma espécie pode cumprir a previsão de uma morfologia inferida?Não é obrigado uma forma intermediária ter apenas uma linha de descendentes, assim é possível detectar em fósseis o padrão de hierarquias aninhadas numa classificação taxonômica, que é a evidência da descendência comum.É por isso que não saber com certeza quem foram os ancestrais (algo dispensável para a Cladística) não é justificativa para se ter a atitude danosa de encorajar os outros a concluir que nunca houveram ancestrais, já que a presença de formas transicionais indica sim que houveram ancestrais.
Prediz-se formas transicionais baseado nos estados derivados compartilhados por pares de táxons (é feita uma matriz dos estados de caracteres compartilhados) e nas mudanças que teriam que ocorrer para transformar um táxon em outro.Um exemplo do primeiro caso, é que dizemos que as aves é a classe que teriam formas intermediárias com os répteis porque a última é a que mais compartilha traços unicamente derivados com elas.E no segundo caso, podemos citar o exemplo em que se esperaria ver caudas longas e dentes na boca em transicionais entre répteis e aves e os transicionais entre animais terrestres e cetáceos deviam ter pélvis desenvolvidas, e é assim que ocorre em vários deles.Formas intermediárias existem até hoje.Por exemplo, répteis são transicionais entre anfíbios e mamíferos.
Como Gee mesmo analisou em In Search of Deep Time ( ver capítulo 1 no site do NY Times- http://www.nytimes.com/) , aproximações de mensurações não existe apenas em biologia evolutiva, mas em todas as ciências.Valores exatos de mensurações quase nunca são sabidos em ciência.Saber todos os ramos de todas árvores filogenéticas de todas as espécies seria um descrição ultra-rigorosa da descendência comum que jamais poderia ser sabida com certeza.O mesmo ocorreria com o conhecimento sobre o valor exato da constante gravitacional universal, já que até este tem medidas independentes que desacordam."Pegar uma linhagem de fósseis e afirmar que eles representam uma linhagem" é uma atitude apenas da velha paleontologia que constrói árvores de ancestrais e descendentes, porque infelizmente ainda existem paleontólogos do elo perdido.Esta corrente perdeu sua força com o surgimento da Cladística, que segundo Gee é a única abordagem da sistemática capaz de produzir hipóteses testáveis sobre a relação entre organismos.Como Gee não reinventou a roda com essas declarações, fica claro porque ele protestou contra essas citações seletivas e fora-de-contexto.Vejam em trecho da resposta dele publicado na NCSE (em "Gee Responds to Discovery Institute Use of Quotations" ):
"O Discovery Institute tem utilizado sem autorização citações seletivas de meu livro In Search of Deep Time para apoiar suas visões obsoletas e equivocadas".
Como as formas transicionais evidencia a evolução obedece a seguinte lógica.Quando uma linhagem se ramifica em duas, uma ou ambas linhagens podem adquirir funções novas. Desde que uma nova linhagem deve recrutar e modificar estruturas pré-existentes para executar estas funções novas, a mesma estrutura compartilhada por estas duas espécies executará agora uma função diferente em cada uma das duas linhagens.Desde que as características derivadas evoluíram de umas estruturas mais primitivas, então é possível fazer predições detalhadas e explícitas sobre as morfologias possíveis de intermediários fósseis.Um exemplo disso é que a transição réptil-mamífero no registro fóssil.Vemos uma ordem fóssil com animais de mandíbulas reptelianas adquirindo cada vez mais características de mandíbulas mamíferas.Muito pelo contrário, o que vemos são répteis adquirindo estruturas precursoras as existentes em mamíferos.Palato secundário primitivo (Procynosuchus), aumento no osso dentário e dentes com múltiplas cúspides (Cygnognathus), junta dupla na mandíbula (Probelosodon).Sobre o último caso, cito um notável paleontólogo:
"O maxilar inferior dos répteis é formado por vários ossos, e o dos mamíferos apenas por um. Os ossos do maxilar inferior dos não-mamíferos foram reduzindo-se pouco a pouco, até se tornarem pequenos vestígios atrofiados, localizados na parte posterior da mandíbula dos mamíferos ancestrais. O martelo e a bigorna do ouvido dos mamíferos descendem dessas atrofias. 'Como se processou uma transição como essa?', os criacionistas perguntam. Por certo, um osso ou está na mandíbula ou encontra-se no ouvido. No entanto, os paleontólogos descobriram duas linhagens de transição entre terápsides (ou assim chamados répteis que parecem mamíferos) com dupla articulação maxilar - uma delas formada pelos antigos ossos quadrático e articular (que logo se transformariam em martelo e bigorna) e a outra formada pelos ossos esquamosais e dentários (como nos mamíferos modernos)."(Gould,1981).
A mesma lógica vemos em outros casos: crânios desenvolvidos de homidídeos aparecem gradualmente a partir de crânios simiescos,nadadeiras de mamíferos aquáticos são antecedidos por patas natatórias, asas são antecedidas por membros longos de ossos pneumáticos.Poderia existir uma forma anatômica nova com toda uma sequência invertida, mas não há. E a evolução também poderia ser falsa se se encontrassem certos fósseis intermediários proibidos como transicionais entre mamíferos e aves, Equus e Homo, Elephantidae e Cetacea.Deixo como um exercício de imaginação para o leitor, os exemplos de muitíssimos outros intermediários que seriam proibidos segundo a teoria da evolução.
Filogenias da Cladística são baseadas na morfologia.A ordem em que os táxons surgem é uma dedução de uma filogenia.A estratigrafia fornece dados independentes para a inferência da ordem do surgimento das formas intermediárias e esses dados podem ser comparados com a ordem predita pela Cladística.A congruência da cronologia dos ancestrais comuns obtidas por dados independentes (filogenia e estratigrafia) já foi corroborada com significância estatística várias vezes na literatura científica (Norell & Novacek,1992;Benton & Storrs,1994).Nos dois papers citados, essa situação é válida para a maioria dos cladogramas analisados.Outra dado é que a "árvore filogenética padrão" deste gráfico do Talk Origins Archive mostra uma ínfima incongruência estatística, com o valor de P (que representa a razão do número máximo de árvores inconsistentes sobre o número total de árvores possíveis) abaixo de 0,0001 (Theobald,2004).Podemos citar também casos de incongruências na cronologia dos ancestrais comuns de filogenias consensuais que não seriam admitidas em nenhuma hipótese e que nunca foram achadas: mamíferos incrustados em rochas do Devoniano e homínideos incrustados em rochas do Jurássico.
Ainda assim, não há nada na história da evolução que diz que uma forma intermediária tem que se extinguir quando evolui uma linha de descendentes.Sobre esse assunto, em Ícones da Evolução Jonathan Wells diz que as formas intermediárias com a morfologia dos ancestrais hipotéticos do Archaeopteryx não aparece até 10 milhões de anos depois do Archaeopteryx.Entretanto, isso é falso e irrelevante.Existem dinossauros terápodos mais antigos que o Archaeopteryx, incluindo o Ornitholestes que viveu entre 156 e 145 milhões de anos e possui uma morfologia levemente mais distante das aves que o Archaeopteryx (Carroll,1997.p.309).Dada a insuficiência do registro geológico é comum que espécies aparecem, desapareçam e reapareçam de novo.Por exemplo, o celacanto não aparece no registro fóssil há 80 milhões, contudo ele está vivo hoje e nem por isso Wells acredita que ele "renasceu" ( e se fizesse isso estaria admitindo que acredita na criação especial).Por isso, os 10 milhões de anos são insignificantes , representando apenas 0,2% do tempo geológico da vida na Terra e o Archaeopteryx não está "fora de lugar" por causa disso, porque a congruência entre esses dados independentes deve ser aproximada e não perfeita.Métodos estatísticos que avaliam as dicrepâncias entre dados independentes, assim como o método mais popular que estima a confiabilidade das árvores, já foram analisadas na parte 2 deste artigo.
PALEONTOLOGIA REFUTA DARWIN?
Comumente, são utilizadas citações de Darwin sobre a relativa ausência de formas transicionais em que ele via como um potencial refutação de sua teoria.Acontece que na época de Darwin, a paleontologia estava apenas engatinhando.Mas será que Darwin viu a abundância de transições de espécies como um teste crucial para a teoria evolutiva?Ele disse:
“O registro geológico é extremamente imperfeito e este fato em grande medida há de explicar porque não se encontramos variedades intermináveis, ligando todas as formas de vida extintas e existentes pelos mais tênues passos graduados. Quem rejeitar estes pontos de vista sobre a natureza do registro geológico rejeitará justificadamente toda minha teoria”(Dawkins,cit.Darwin,2001.p.353).
Veja que mesmo Darwin foi cauteloso afirmando que esta poderia não ser uma objeção fatal e isso mostra que ele não tinha informação suficiente para retrodizer as condições iniciais, então muito cuidado em utilizar o “falseacionismo ingênuo” como método científico.Analogamente, não é porque a observação de dois corpos de massas diferentes caindo a mesma velocidade seja um confirmador de uma teoria da física galileana, signifique que o contrário a torne falsa.Senão, vejamos.Será que há motivo para se rejeitar a natureza do registro fóssil que Darwin tanto fala? Temos apenas 300 mil espécies registrados em fósseis, um número que é uma ínfima parte até do número de espécies viventes, que é estimado entre 10 e 50 milhões (Purves et al. 2002.p.384).E o número baixo de fósseis nada tem haver com criação recente da Terra ou outra coisa, apenas a raridade do fenômeno da fossilização.Ninguém ignora hoje que a fossilização só preserva um animal se ele morrer num ambiente com muitas condições favoráveis a fossilização (presença de sedimentação favorece) e ainda por cima, esses sedimentos devem se compactar em rochas e não sofrerem erosão por milhares de anos.Assim, o número baixo de fósseis nada tem haver com criação recente da Terra ou outra coisa, apenas a raridade do fenômeno da fossilização.É falsa a afirmação que há poucos transicionais, porque as transições não devem existir apenas se for entre espécies.Só na transição entre monos e Homo sapiens temos além do Ardipithecus ramidus, os Australopithecus anamensis, afarensis, africanus, aethiopicus, boisei, robustus, garhi e os Homo habilis, georgicus, rudolfensis, erectus, ergaster, antecessor e heildelbergensis.Como diria um suposto oponente de Darwin:
"As formas transitórias geralmente não existem no nível das espécies, mas são abundantes entre os grupos mais amplos"(Gould,1981).
Mesmo o fato descrito por Gould não impede que tenhamos a evidência observacional de uma espécie se transformando gradualmente em outra, coisa possível devido ao fenômeno "espécies-em-anel" (ver parte 2 deste artigo).
E porque as transições de espécies são tão raras?Na verdade existem não existem motivos para elas serem abundantes , como ajudaram a esclarecer os proponentes da Teoria do Equilíbrio Pontuado, Stephen Jay Gould e Niles Eldredge.Os dois cientistas analisaram que o registro fóssil não é só imperfeito, mas imperfeito justo quando a mudança evolutiva está ocorrendo.A questão é que mesmo que registro geológico fosse perfeito, não encontraríamos essas transições ligando as espécies em pequenos passos se escavássemos um só local pelo simples fato que a maior parte da evolução se dá em locais diferentes de onde encontramos os fósseis, já que as condições para especiação e redução de fluxo gênico só podem ser comum em isolamento geográfico.Assim, novas formas que evoluíram em outras áreas mais cedo ou mais tarde invadirão a área da sua espécie ancestral. Dessa forma, a maioria dos locais em que fósseis são encontrados, a transição de uma espécie em outra será abrupta. Essa mudança abrupta irá refletir a substituição por migração de qualquer forma, não a evolução.O fato das extinções serem a regra ao longo da história da vida também contribui para isso; um princípio ecológico amplamente aceito diz que duas espécies aparentadas de modo de vida semelhante não podem coexistir porque elas entram em competição (Dawkins,2001.p.350).Assim, para encontrar-se esses tipos de formas transicionais, a área de especiação precisa ser encontrada. Quando grandes áreas são pesquisadas, formas transicionais que fecham a lacuna entre as duas espécies podem ser encontradas em pequenas áreas localizadas, caso a espécie não ocupe tão breve intervalo de tempo que precisássemos de um registro fóssil extremamente rico para reconstituí-la.Stephen Jay Gould e Niles Eldredge conseguiram descrever uma destas áreas - uma pequena pedreira isolada em Nova York que ilustrou a transição de uma espécie de trilobita Phacopsto para outra, os níveis mais baixos continham as espécies originais de trilobitas, os níveis superiores continham as espécies novas e no meio havia uma série de transições.Em outros locais, a transição era abrupta.
Assim, a Teoria do Equilíbrio Pontuado não é uma proposição ad hoc: refletem predições da teoria evolucionista, e não imperfeições de fósseis.Ela é uma teoria baseada numa evidência positiva e baseado em estudos de grupos de espécies vivas e extintas (Eldredge & Gould, 1972). Por fim, a moderna teoria genética sustenta que a alteração moderada num caráter exigirá pelo menos 50 mil gerações, coisa demonstrada em análise de registros fósseis mais contínuos ; uma mudança em um caráter que consuma um período de tempo bem maior deverá requerer seleção tão fraca que esta será superada pela deriva genética (Futuyma,1992.p.425,426).Então, a própria genética de populações prediz que a evolução por seleção natural será pontual e que as transições fósseis de espécies serão raras, embora o mesmo não possa ser dito para grupos mais amplos.
Muita gente confunde a estase postulada pelos proponentes do Equilíbrio Pontuado com a descontinuidade do registro fóssil.Nenhum proponente do Equilíbrio Pontuado postula que a ausência mutações direcionais em infinitesimais passos no registro fóssil, é a prova de um padrão de evolução morfológica que flutua pouco e rapidamente de forma cegamente rápida.Nenhum biólogo sério, como o Niles Eldredge, acredita que uma linhagem de Australopithecus passou bom tempo sem mudança morfológica e de repente deu origem a um filho mutante que tinha um crânio tão diferente de seu pai que deu origem a uma espécie de outro gênero (neste caso específico, o gênero Homo).A estase real só ocorre uma vez que as espécies foram originadas e estão adaptadas ao novo nicho ecológico em que se encontram, e tendem a permanecer como estão por um bom tempo geológico.Para explicar a estase, pontuacionistas se baseiam numa idéia da genética de populações que diz que uma população ampla tende a ter dificuldades de sair da inércia genética, explicando a estase em populações com tempo superior a milhões de anos.E que se a maioria das especiações ocorre como Mayr imaginou, então o efeito desestabilizador do pequeno tamanho populacional (efeito fundador) é eficiente para mudar populações em direção a novos picos adaptativos.Apenas em certas situações esse argumento não é válido, segundo geneticistas de populações, que argumentam que o fluxo gênico não é eficiente para se contrapor a seleção forte.Isso não quer dizer que longos períodos sem grandes mudanças morfológicas seja contraditório com a evolução.Qualquer evolucionista concorda que as taxas de evolução variam e que ausência de pressão seletiva provoca estase morfológica.Não há absolutamente nenhum mistério nisso, porque a seleção estabilizadora também é uma força da seleção natural.Mas existem alguns casos de estase em casos de instabilidade ambiental.Como explicá-la?A primeira explicação é extrínseca.Os organismos não são alvos passivos de influências ambientais, mudam de habitat, definem e criam os ambientes em que vivem.Observa-se que muitos animais que tem capacidade de dispersão podem escolher os hábitats em que eles vivem ou estes permanecem constante, apesar do ambiente mudar ao longo do tempo (Futuyma,1992.p.427).A alteração de preferência de localização pode explicar as explosões repentinas.Se esta for a maior parte causa da estase, então a seleção estabilizadora também é capaz de explicá-la.Outro explicação é limitação interna sobre modificações evolutivas, onde alguns traços são mais difíceis de serem alterados.O último caso não tem evidência de dados observacionais,porque criadores de animais que fazem cruzamentos seletivos não vêem indícios forças antievolutivas intrísecas (Dawkins,2001)
Muito se fala da diferença entre o neodarwinismo ortodoxo e o Equilíbrio Pontuado, mas a diferença entre essas teorias é menor do que se poderia imaginar.Numa árvore de Equilíbrio Pontuado representando a diversificação morfológica dos táxons num contexto de curto prazo geológico, geralmente as mudanças ocorrem mais rapidamente e apenas próximas dos eventos de especiação.No caso do Gradualismo Filético, geralmente a mudança ocorre mais lentamente e não se acentua durante os eventos de especiação.Mas num contexto de um prazo bem maior que os eventos de especiações, uma árvore desenhada por pontuacionistas representando a diversificação morfológica dos táxons ao longo do tempo, ficaria quase igual à desenhada pelos neodarwinistas ortodoxos; porque os últimos defendem que quando as mudanças substanciais ocorrem, são rápidas de um modo que fazem o registro fóssil parecer pontuado, com a diferença que não há uma conexão necessária entre especiação e mudança morfológica (Futuyma,1992.p.424).Os pontuacionistas são defensores da hipótese da velocidade continuamente variável, ou seja, para eles a velocidade ou é rápida ou nula (estase), com a ausência quase completa de ritmos intermediários.Outros neodarwinistas são defensores da hipótese da velocidade continuamente variável.Não vêem necessidade de salientar certas velocidades mais que outras, porque defendem que a velocidade de evolução varia continuamente de muito lento a muito rápido, passando por ritmos intermediários.Qualquer pessoa bem informada pode ver que é falso dizer que a teoria do Equilíbrio Pontuado incompatível com o que Darwin imaginou como gradualismo.Tanto Equilíbrio Pontuado quanto Gradualismo Filético são fatos reais acerca do registro fóssil.Elas não são escolas de pensamento advogadas por lados opostos.As duas teorias apresentam pontos extremos de duas dimensões contínuas sobre as taxas de evolução, de tal modo que é até possível dúvida se algumas sequências fosselíferas mostram gradualismo ou estase com pontuação.Isso porque antes de se certificar com certeza se um padrão é gradual ou pontuado, deve-se ter uma sequência fosselífera relativamente completa (Ridley,2006.p.622).
E o fato que as taxas de evolução podem variar é conhecido desde o século XIX.Em A Origem das Espécies , Darwin diz que “é provável que os períodos, durante os quais as espécies sofreram modificações, embora muitas e por muito tempo (medido por anos), têm sido curtos em comparação com os períodos durante que cada um permaneceu em uma condição inalterada” (final da 6ª edição, 1872).Em nenhum momento em A Origem Das Espécies, Darwin fala em “mudança constante” ou "velocidade constante" da evolução, seu gradualismo apenas diz que não podem ocorrer saltos numa única geração.Nem mesmo as “pontuações” são observações antidarwinistas.Mudanças morfológicas podem ocorrer rapidamente, geologicamente falando, e ainda assim serem geneticamente graduais.
Ainda assim, algumas críticas são feitas a pontuacionistas e elas atingem mais a parte da teoria que postula uma conexão entre alteração morfológica e especiação.Por exemplo, seus críticos acreditam que a estase considerada pelos pontuacionistas é puramente morfológica.Argumentam que não é possível delimitar as espécies pelo modelo fenético, ou seja, pela aparência.Há grande número de espécies que não são distinguidas pela morfologia, mas constituem unidades evolutivas diferenciadas, inclusive isoladas reprodutivamente, mesmo quando coabitam o mesmo nicho. Mesmo na estabilidade da morfologia, as populações podem evoluir e se especiar: as taxas evolutivas dos genes e as da morfologia não estão pareadas.A constância morfológica pode acompanhar grandes modificações genéticas (Stearns & Hoekstra,2003.p.68 e 124).Além disso, ao contrário do que pontuacionistas pensam, a especiação não parece ser um requerimento para grandes mudanças morfológicas.Há grandes polimorfismos dentro de algumas espécies. Por exemplo, os peixes guelra-azul têm duas formas de macho com morfologias um tanto distintas.Para que se confirme que mudança rápida é acompanhada de especiação, seria necessário mostrar que uma forma ancestral não alterada persiste simpatricamente com seu descendente modificado, coisa que tem poucos casos documentados.
Mas apesar de tudo, é um argumento simplista antievolucionista afirmar que Gould ou Elderdge postularam que o registro fóssil não pode "comprovar o gradualismo".O gradualismo não é uma afirmação quanto a taxas ou tempo da evolução (Mayr,2005.p.122).Eventos geneticamente graduais são mudanças dentro da área de variação biológica esperada entre duas gerações consecutivas.Assim, o gradualismo requer apenas restrição a possíveis mecanismos de descendência comum e adaptação, não afirmando nada sobre se a evolução deve ser constante,rápida, ocorrer frequentemente,etc.Um exemplo ajuda a nos entender isso.O palentólogo George L. Stebbins criou uma hipótese matemática segundo a qual é possível uma espécie de 40 gramas evoluir por 12 mil gerações (cerca de 60 mil anos) até chegar o peso de um elefante (6 toneladas), e ainda assim o fenômeno ser geneticamente gradual e ser imperceptível em termos de tempo da vida humana.Considerando que o tamanho corpóreo fosse o único fator que se alterasse nessa evolução, teríamos um caso hipotético de uma aparição extremamente abrupta no registro fóssil, mas que ainda assim não refutaria o gradualismo darwiniano.Antes de dizer que o registro fóssil contradiz o gradualismo é necessário mostrar que o tempo que separa um organismo que aparece abruptamente no registro fóssil e seu antecessor intermediário mais cladisticamente próximo, indique que o tempo que os separam funcione como uma restrição a possíveis mecanismos de descendência comum e adaptação.Uma maneira de se medir a taxa de evolução de caracteres é o "darwin", que representa uma mudança de um fator de e (a base dos logaritmos naturais, 2,718) por milhões de anos.Dado que a taxa morfológica média de eventos de colonização é de 400 darwins - suficiente para transformar um rato num elefante em 10.000 anos(Gingerich ,1983)- a evolução de um gênero que requeresse uma taxa morfológica maior que a média da dos experimentos de seleção artificial (58.700 darwins) ou muito superior a dos eventos de colonização (digamos, 4.000 darwins) seria uma objeção fatal a evolução.Entretanto, a taxa morfológica do registro fóssil mais rápida dos mamíferos pós-pleistocênicos que Gingerich analisou foi de 32 darwins!
Consequentemente, deve-se saber que quando nos referirmos a gradualismo filético, trata-se de uma hipótese sobre o padrão de macroevolução e da distribuição das mudanças morfológicas ao longo da longevidade filética das espécies e que foi formulado por Ernst Mayr, não por Darwin.De fato, biólogos pontuacionistas também são gradualistas darwinianos, porque não dão importância significativa aos saltos macromutacionais na evolução.Somando todas essas observações, podemos afirmar com segurança que a evidência fóssil não é anti-gradualista ,e consequentemente, anti-darwinista.
Referências:
Benton, M. J., and Storrs, G. W. (1994).Testing the quality of the fossil record: paleontological knowledge is improving. Geology 22: 111-114.
Carroll, R. L. (1997). Patterns and Processes of Vertebrate Evolution. Cambridge: Cambridge University Press.
Darwin, C. (1987).A Origem das Espécies.Editora Ediouro, Rio de Janeiro-RJ.
Dawkins, R. (2001).O Relojoeiro Cego: A Teoria da Evolução contra o Desígnio Divino.Editora Companhia das Letras, São Paulo-SP.
Eldredge, N. and Gould, S. J. (1972). Punctuated equilibria: An alternative to phyletic gradualism. In: Models In Paleobiology (T. J. M. Schopf, ed.), San Francisco: Freeman, Cooper & Co., 82-115.
Futuyma, D. J. (1992). Biologia Evolutiva, 2ª edição.Sociedade Brasileira de Genética & CNPq, Ribeirão Preto-SP.
Gingerich, P. D. (1983).Rates of evolution: Effects of time and temporal scaling.Science 222: 159-161.
Gould, S.J. (1981).Evolution as Fact ant Theory. Discover.
Dawkins, R. (2001).O Relojoeiro Cego: A Teoria da Evolução contra o Desígnio Divino.Editora Companhia das Letras, São Paulo-SP.
Mayr, E. (2005).Biologia, Ciência Única.Editora Companhia das Letras, São Paulo-SP.
Norell, M. A. and Novacek, M. J. (1992).The fossil record and evolution: Comparing cladistic and paleontologic evidence for vertebrate history. Science 255: 1690-93.
Purves, W.K., Sadava, D., Orians G.H., Craig Heller, H. (2002).Vida: A Ciência da Biologia, 6ª Edição. Editora Artmed, Porto Alegre-RS.
Ridley, M. (2006).Evolução, 3 ª edição, Editora Artmed, Porto Alegre-RS.
Stearns S.C. e Hoekstra, R.F. (2003).Evolução: Uma Introdução.Atheneu Editora São Paulo, São Paulo-SP.
Theobald, Douglas L. "29+ Evidences for Macroevolution: The Scientific Case for Common Descent." The Talk.Origins Archive. Vers. 2.83. 2004. 12 Jan, 2004
posted by espectro109 at 6:44 PM
22 comments
![]()

{kind=link}
{kind=link}
{kind=link}